A comprehensive review of siRNAs and shRNAs as tools for gene silencing.
RNA interference (RNAi) is the process by which the expression of a target gene is effectively silenced or knocked down by the selective inactivation of its corresponding mRNA by double-stranded RNA (dsRNA). RNAi is activated by dsRNA species delivered to the cytoplasm of cells. The silencing mechanisms can either lead to the degradation of a target mRNA, as induced by small interfering RNAs (siRNAs) or short hairpin RNAs (shRNAs), or the suppression of translation of specific mRNAs, as induced by microRNA (miRNA). The focus of this review will be how shRNAs and siRNAs lead to protein knockdown. Through the activity of several proteins (discussed below), targeting of a cellular mRNA by short, anti-sense nucleic acids (siRNAs and shRNAs) results in its subsequent degradation. This, in turn, blocks further expression/accumulation of the proteins, leading to a decrease in its levels, and eventual knockdown.

As early as 1984, it was observed that anti-sense RNA was capable of inhibiting gene expression. In 1993, Nellen and Lichtenstein proposed a model to explain this observation. However, it was not until 1998 that Fire et al published results on RNAi in Caenorhabditis elegans identifying dsRNA as being substantially more effective at inhibiting gene expression than single-stranded RNA. It was eventually determined that the miRNA pathway involved many of the same protein components as the RNAi pathway.
The key components of the RNAi machinery are summarized in Table 1. They include the dsRNA targeting the target gene(s) (either siRNA or shRNA), Dicer, the Argonaute family of proteins (specifically Ago-2), Drosha, RISC, TRBP, and PACT. Drosha and Dicer also possess other non-canonical functions [2].
| Term | Description |
|---|---|
| siRNA | Small interfering RNA (siRNA). dsRNA with 2 nt 3’ end overhangs that activate RNAi, leading to the degradation of mRNAs in a sequence-specific manner dependent upon complementary binding of the target mRNA. |
| shRNA | Short hairpin RNA (shRNA) that contains a loop structure that is processed to siRNA and also leads to the degradation of mRNAs in a sequence-specific manner dependent upon complementary binding of the target mRNA. |
| Drosha | An RNase III enzyme that processes pri-miRNAs and shRNAs in the nucleus. |
| Dicer | Ribonuclease (RNase) III enzyme which processes dsRNAs into 20–25 bp siRNAs leaving a 2 nt overhangs at the 3’ end. Drosophila Dicer-2 cleaves long dsRNAs, while Dicer-1 is important for miRNA processing. |
| RISC | The minimal RNA-induced silencing complex (RISC) consists of the Argonaute protein and an associated siRNA. It may also contain PACT, TRBP, and Dicer. It should be noted that the exact composition of RISC has yet to be described. |
| TRBP | Needed for dsRNA cleavage by Dicer and subsequent passage to the RISC. |
| PACT | Protein R (PKR)-activating protein (PACT). Associates with Dicer and TRBP for dsRNA cleavage. |
| Argonaute family of proteins | Along with the single-stranded siRNA, these assemble to form the RISC. Bind 21–35 nt RNAs including miRNAs and siRNAs, and their associated target mRNA and then cleaves them through its endonucleolytic function. Cleaves between the 10th and 11th nucleotides of the anti-sense (or guide) RNA. |
Two key approaches to RNAi that have gained substantial interest for use in gene silencing are the double-stranded small interfering RNAs (siRNAs) and the vector-based short hairpin RNAs (shRNAs). While both siRNAs and shRNAs (Figure 1) can be used for protein knockdown, there are differences in their mechanisms of action (Figure 2). Either long dsRNAs or short duplexes of about 21 base pairs (bps) can be introduced directly into cells in tissue culture (see Mechanisms of Delivery for further details). While there are a few reports of siRNAs being translocated to the nucleus upon transfection into cells, it is more generally accepted that they accumulate in the cytoplasm. Upon introduction to the cell, the long dsRNAs form a complex with Dicer [3], a dsRNA-specific RNase III enzyme that processes them into 21-23 nucleotide (nt) siRNAs with characteristic 2 nt 3’ overhangs. The cleaved products are then incorporated into the RISC, which is composed of Argonaute-2 (Ago-2), Dicer, and TAR-RNA-binding protein (TRBP). The RNA duplex is separated, and one strand is removed from the complex. The strand with the lowest duplex stability at its 5'-end is selected for stable incorporation into the RISC.
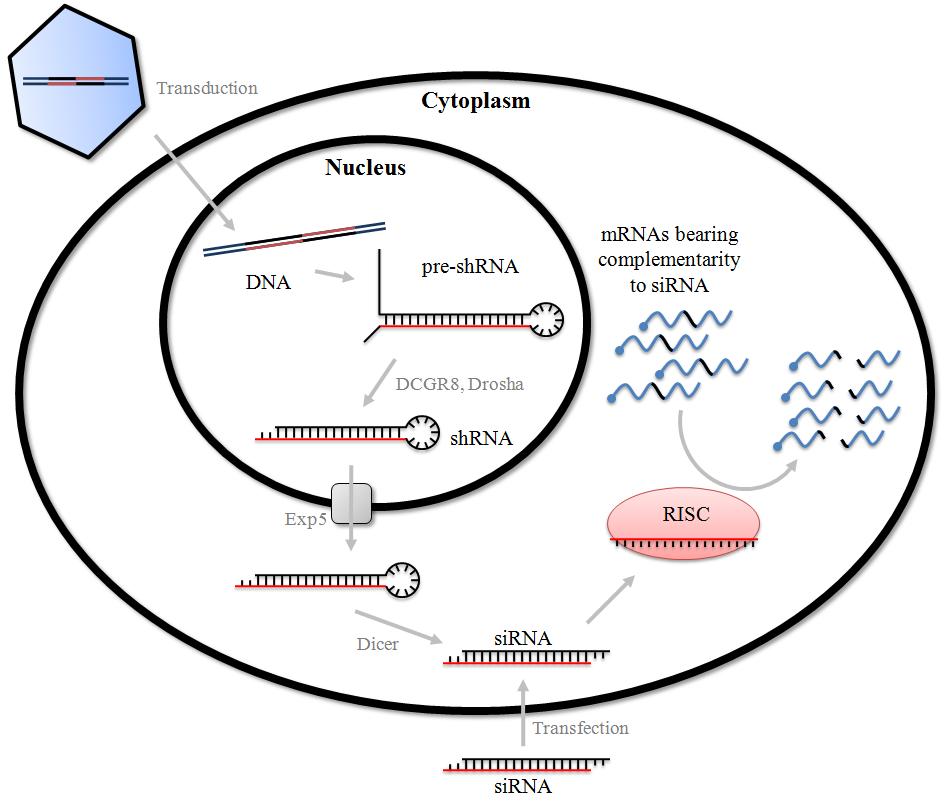
shRNAs are synthesized in the nucleus of transfected/transduced cells and form hairpin structures that consist of a stem region of paired antisense and sense strands connected by unpaired nucleotides that make up a loop (Figure 1b and 1c). They are converted into siRNAs by the same RNAi machinery that processes miRNAs. shRNAs are introduced into the nuclei of target cells using either bacterial or viral vectors that, in some cases, can stably integrate into the genome. shRNAs are transcribed by either RNA polymerase II or III, depending on the promoter driving their expression. These initial precursors are processed by Drosha and its dsRNA-binding partner DGCR8, resulting in species known as pre-shRNAs, before being exported to the cytoplasm by Exportin-5. The pre-shRNA is then cleaved by Dicer and TRBP/PACT, removing the hairpin and creating a 20–25 nt double-stranded siRNA with 2 nt 3’ overhangs at each end. This active siRNA is then loaded onto the RISC complex.
Once loaded onto the RISC, the process of target mRNA recognition and degradation by both shRNA and siRNA is essentially the same. As a moiety of the RISC, the siRNA binds to the target mRNA in a sequence-specific manner that is mediated by complementary base pairing, leading to cleavage of the target RNA phosphate backbone near the center of the duplex via the action of the RNase-H like activity of Ago-2. An interesting feature of this system in some organisms is that annealing of the siRNA to the target mRNA allows the siRNA to act as a primer, while the target mRNA acts as a template for an RNA-dependent RNA-polymerase. This creates a new dsRNA, which is then processed by Dicer, creating a positive feedback loop that increases the pool of siRNAs. It should be noted that siRNAs typically require perfect homology to induce degradation. This process is illustrated in Figure 2.
The process by which the target mRNA is found by the RISC is not well understood. However, a report by Ameres et al revealed that the accessibility of the target sequence of the cellular mRNA affected its cleavage. They also noted that the RISC is not capable of unfolding RNAs. They proposed a model in which the RISC comes in contact with ssRNAs in a nonspecific manner by random diffusion, with the 5’ end undergoing base-pairing more efficiently than the 3’ end. This appears to determine the stable association between the RISC and the target mRNA.
Advantages of shRNA over siRNA include the ability to use viral vectors for delivery to overcome the difficulty of transfecting certain cell types, the option to control shRNA expression using inducible promoters, and the ability to co-express them with a reporter gene. Additionally, they may cause fewer off-target effects (discussed further below).
Many labs have published strategies for the creation of long dsRNAs for transfection experiments. However, the efficacy of each approach is dependent on the system. Also, long dsRNAs activate the innate immune response, leading to cell death. Factors that affect the activity of shRNAs include loop structure, thermodynamic properties of the hairpin, secondary structure, and the surrounding sequences. When choosing between siRNAs or shRNAs, an important factor to consider is the length of the experiment. siRNAs are transiently expressed in cells, while shRNAs can be stably integrated through virus-mediated transduction.
Guidelines for siRNA/shRNA design are available from major manufacturers of RNAi products.
- siRNA sequences between 19–29 nt are generally the most effective. Sequences longer than 30 nt can result in nonspecific silencing.
- Ideal sites to target include AA dinucleotides and the 19 nt 3’ of them in the target mRNA sequence. Typically, siRNAs with 3’ dUdU or dTdT dinucleotide overhangs are more effective. More recent data indicates that other dinucleotide overhangs maintain activity, however siRNAs can be cleaved by RNase at single-stranded G residues and therefore GG overhangs should be avoided.
- Choose at least 2–4 target sequences at different positions since mRNAs tend to be highly structured and bound by regulatory proteins. Avoid designing siRNAs with a 4–6 poly(T) tract, since it acts as a termination signal for RNA pol III.
- Check to make sure your siRNA sequences do not have homology to other coding sequences.
- Sequences should have a G/C content between 35–55%.
| Company/group | Program Name | Description | Sample Reference |
|---|---|---|---|
| Thermo Scientific | siDESIGN | User-friendly siRNA design tool. Allows you to choose the region the siRNA targets (5’ or 3’ UTR or ORF), G/C percentage, and if you want to BLAST search the sequence. | |
| Naito et al | siDirect | Identifies siRNA targets based on nucleotide sequence. Provides location within the sequence, melting temperature of seed duplex, and a minimum number of mismatches against off-target sequences. | |
| Invitrogen | BLOCK-IT RNAi Designer | Identifies siRNA, shRNA, and miRNA targets within nucleotide sequences. Also converts siRNA sequences into shRNA sequences. | |
| InvivoGen | siRNA Wizard | Searches a coding sequence for siRNAs, designs scrambled sequences and hairpin inserts based on your siRNA sequence. | |
| Gene Link | shRNA design tool | shRNA design tool that allows you to choose between three loop sequences or enter a custom sequence, as well as specify restriction sites for the 5’ and 3’ ends, designate GC content, and length. | |
| transomic | shRNA design tool | shERWOOD-UltramiR shRNAs are "based on the functional testing of over 250,000 shRNA sequences using a high-throughput sensor assay (Knott et al 2014) and use key sequence characteristics for predicting shRNA potency to select the rare shRNA designs that are potent at single copy representation in the genome." | [4, 5] |
shRNAs should consist of the sense and antisense sequences (each 19–21 nt in length) separated by a loop structure, and a 5’ AAAA overhang. When designing the loop structure, Ambion scientists and others recommend using a 9 nt spacer (TTCAAGAGA), while Invivogen uses a 7 nt loop (TCAAGAG) for certain vectors, though this can vary depending on your system. Loop sequences of 3 to 9 nt in length have been shown to be effective. Additionally, when creating the shRNA cassette, they suggest that the sense strand come first, followed by the spacer and then the antisense strand. 5’ overhangs in the shRNA construct should be avoided, as they could result in silencing of the shRNA. In addition to manually designing an siRNA or shRNA, there are also several design programs available. A few of them are included in Table 2. Additionally, there are multiple companies that offer premade siRNAs and shRNAs (representative examples are shown in Table 3).
Most shRNAs are transcribed from vectors. Their expression is most commonly driven by either the Pol III U6 promoter, which drives high levels of constitutive expression, or the weaker H1 promoter. A major advantage of shRNA over siRNA systems is that shRNAs can be designed to be inducible. There are both Tet-on and Tet-off inducible systems commercially available, as well as constructs containing a modified U6 promoter that is induced by the insect steroid molting hormone ecdysone. A Cre-Lox recombination system has been used to achieve controlled expression in mice. There are also synthetic shRNAs available which, unlike the viral vector-delivered molecules, can be chemically modified to affect their activity and stability as described above for siRNA molecules.
| Company/Group | Name of Program | Description of what they offer |
|---|---|---|
| Thermo Fisher Scientific | Silencer [6] | Available for all human, mouse, and rat gene targets in the RefSeq database |
| The Broad Institute | Genetic Perturbation Platform [7] | Public-private effort with the mission to create a validated shRNA library and related tools that can be used to determine the function of human and mouse genes. |
| MilliporeSigma | MISSION [8, 9] | Offers shRNAs as bacterial glycerol stocks, plasmid DNA, or lentiviral particles |
| Horizon Dharmacon | SMARTchoice SMARTpool [10] Accell [11] ON TARGETplus siRNA [12] | Offers shRNAs as lentiviruses, among others |
| Cellecta | DECIPHER [13] | Pooled libraries for hiigh-throughput screening |
To ensure that the effects observed after RNAi treatment are the result of gene silencing and not merely the introduction of the siRNA/shRNA or from activation of RNAi pathways, it is important to include the appropriate controls (Table 4). The two most common controls are scrambled and non-targeting controls. A scrambled control is exactly what it sounds like, it involves taking the siRNA or shRNA sequence and randomly rearranging its nucleotide sequence. AddGene plasmid 1864 pLKO.1 scramble control shRNA from D. Sabatini at Whitehead Institute, Cambridge, MA is the most used control. A non-targeting control, on the other hand, is an siRNA/shRNA sequence designed such that it does not target any known genes in the target organism. These controls activate the RNAi machinery and allow baseline determination of the effect of the introduction of duplex RNA on gene expression. However, it should be noted that even non-targeting siRNA controls induce a stress response within cells. While both of these types of control sequences will be incorporated into Dicer and activate the RNAi pathway, it is possible that the scrambled control may target an unintended mRNA. As such, care should be taken in designing the scramble sequence to make sure it follows the guidelines mentioned above, and does not target another mRNA sequence.
For shRNAs other important controls include an empty vector control, which contains no shRNA insert, allowing for the determination of the effects of transfection/transduction on gene expression and the response of the cell. Finally, untreated cells (no transfection or transduction) provide a reference for the treated cells and will allow you to determine the cytotoxicity of a particular siRNA delivery method.
| Company/Group | Description |
|---|---|
| MilliporeSigma | shRNA non-targeting control (SHC002 [14] ) , siRNA non-targeting controls (e.g., SIC001 [15] ) |
| Empty Vector Control | |
| ThermoFisher / Invitrogen | shRNA positive control (targets eGFP) |
| Empty Vector control | |
| Scrambled siRNA control | |
| Actin, GAPDH, or GFP positive control | |
| QIAGEN | Scrambled siRNA duplexes [16, 17] |
The exact protocol for delivery of the siRNA or shRNA will depend on the cell type you are working with, since different cell types have varying sensitivities to the introduction of nucleic acids, and whether you are using siRNA- or shRNA-mediated knockdown, as well as the length of the assay you are performing. Transfection, electroporation, and certain viral delivery methods are transient, while lentiviral or retroviral transduction stably integrates the shRNA into the cell’s genome, allowing for persistent expression. Transposon-based vectors, such as sleeping beauty [18] or piggyBac [19], have also been used. Table 5 summarizes the main features of different delivery methods.
Among the most common nucleic acid delivery methods are transfection and electroporation. Transfection involves the formation of complexes of nucleic acids with carrier molecules that allow them to pass through the cell membrane. siRNAs, and sometimes an shRNA encoded on a plasmid, are usually introduced to cells using these methods. Transfection reagents can either be acquired commercially or produced within the lab.
- Lipid Transfection. Cationic lipids that have long hydrophobic chains with positively charged head groups interact with the negatively charged siRNA, surrounding it in a lipid bilayer, which is then endocytosed by the cell.
- Cationic polymer-based nanoparticles. These allow for reduced toxicity and increased efficiency, as well as allowing for the delivery of modified siRNAs.
- Lipid or cell-penetrating peptide (CPP) conjugation. This involves conjugation of the siRNA with a hydrophobic moiety (e.g., cholesterol) or a cationic CCP (e.g., transportin or pentatratin), which promotes delivery into the target cells.
In electroporation (for example, a single 1500 V, 30 ms pulse in a mouse macrophage suspension for 10 pM siRNA [20] ), an electrical field is applied to the cell membrane, which is made up of phospholipid molecules with negatively charged head groups. The electrical pulse causes the phospholipids to reorient, creating pores in the membrane, allowing siRNAs to enter. Electroporation is commonly used for cells that are difficult to transfect. However, the specific settings (voltage, number of pulses, and length of the pulses) must be optimized for each cell type.
- Lentiviruses/Retroviruses
There are many lentiviral and retroviral plasmids that are suitable for shRNA expression (see Products Section below). While the specific plasmid and shRNA design may vary slightly depending on the experiment (e.g., target cell type, etc.), the basic shRNA-expressing lentivirus construct contains the Pol III promoter followed by the shRNA (sense, loop, and antisense sequences followed by five Ts), associated enhancer elements, a 5’ and 3’ LTR, and a packaging sequence. This lentiviral or retroviral plasmid is cotransfected into a packaging cell line (such as 293Ts) along with packaging plasmids, which encode the enzymatic and packaging proteins required for the production of the lentivirus. The replication deficient lentivirus that is produced can be used to transduce the target cells. For detailed protocols for nucleic acid delivery by lentiviruses and retroviruses, please see Nucleic Acid Delivery: Lentiviral and Retroviral Vectors.
- Adenoviral vectors
Adenoviruses are small dsDNA viruses that infect most cell types. Recombinant adenoviruses have had several genes essential for replication deleted, allowing for replication and propagation only in complementing packaging cells (293 cells). DNA delivered using adenovirus-derived vectors are maintained as epichromosomes in the nucleus, making the expression transient, but eliminating the risk of insertional mutations. Adenovirus vectors exhibit extremely broad cell tropism and are safe to work with in laboratory settings. While this is advantageous for experiments in tissue culture, these vectors pose problems in clinical settings. For a more detailed description, please see “ Recombinant Adenoviral Vector Systems”.
- Adeno-associated virus (AAV)
Adeno-associated virus (AAV) is a small, single-stranded DNA (ssDNA) virus that is incapable of replicating without the aid of a co-infecting helper virus, such as herpes simplex virus or adenovirus. AAV infects many cell types, entering the nucleus and undergoing either a lytic (in the presence of a helper virus) or lysogenic (in the absence of a helper virus) life cycle. During infection in the absence of a helper virus, AAV is integrated into the host cell chromosome, resulting in latent infection. Upon infection with a helper virus, the AAV lifecycle is activated and a new virus is produced. AAV2 is the typical serotype used to make recombinant AAV vectors. For example, AR Palla et al delivered shRNA directed against Hpgd in an AAV9 vector [21].
To prevent lytic replication by recombinant AAV particles, they are produced in a similar manner to lentiviral and retroviral vectors: producer cells are transfected with a plasmid that encodes a cloned minimal rAAV genome expressing the desired transgene or shRNA, another plasmid that expresses the AAV genes necessary for the production of an infectious particle, and a third plasmid that encodes the helper genes from adenovirus. AAV vectors provide prolonged and stable expression. It is thought to persist in target cells either in circular form or as linear concatemers. A major drawback to using these viruses is that gene transfer requires relatively high multiplicities of infection, and high titers of recombinant AAV are difficult to produce. Additionally, despite being promising as a viral vector, there are still gaps in our understanding of the AAV life cycle.
| Method | Efficiency of gene delivery | Dividing Cells | Non-dividing Cells | Integration | Cytoplasm or Nucleus | Safety and Experimental Considerations |
|---|---|---|---|---|---|---|
| Transfection | Dependent on cell type, health, and confluency, as well as DNA quantity | Yes | Low efficiency | No | RNA-Cytoplasm Plasmid-Nucleus | Reproducible. Low efficiency in primary cells, nondividing cells and in vivo. |
| Electroporation | High (though this depends on cell types) | Yes | Yes | No | Nucleus and cytoplasm | Good for use on difficult to transfect cells. High cell death and substantial optimization required. |
| Recombinant Adenovirus | 100% | Yes | Yes | No | Cytoplasm | Infect most cell types. Replication-deficient. Capable of handling large (8 kb) inserts. Labor-intensive. |
| Lentivirus | <30% | Yes | Yes | Yes | Nucleus | Minimal chance of mutations resulting from integration. Can infect many cell types. Engineered to infect human cells, so they should be handled with care. |
| Retrovirus | <30% | Yes | No | Yes | Nucleus | Chance of mutations resulting from integration. Engineered to infect human cells, so they should be handled with care. |
| Adeno-Associate Virus | low | Yes | Yes | No | Nucleus | Non-pathogenic. High titers needed |
A limitation of RNAi is that it may not be suited for use in all cell types. For example, the presence of dsRNA-specific RNAses makes them only slightly effective in the neurons of C. elegans. Additionally, delivery of the siRNA or shRNA may not be possible in certain cell types. Some cells do not tolerate transfection and they may not be susceptible for delivery by viral vectors.
An important factor in choosing between siRNA- and shRNA-mediated gene silencing methods is the length of both the assay and the half-life of the target protein. While there have been no large-scale studies comparing the duration and level of protein knockdown achieved by siRNA and shRNA constructs, initial reports using transfected siRNAs and plasmids expressing shRNAs indicated that shRNAs are superior with respect to these criteria. For longer assays, or when trying to knockdown the expression of proteins that have long half-lives, such as p300 (10–22 hours depending on the cell type and conditions ), stable expression of an shRNA may be required.
Protein levels must be assessed to determine the degree and kinetics of knockdown. The most straightforward method of determining if silencing worked is to perform a western blot. Briefly, this involves collecting the treated cells (both RNAi expressing and various controls), lysing them, quantifying the amount of protein, and running the samples on a denaturing SDS-PAGE gel. After detection with a specific antibody, the levels of protein in the RNAi-treated cells can be compared to the controls and knockdown efficiency can be determined. A detailed explanation of western blotting and protein quantitation can be found at Labome.
If inadequate or no knockdown is observed, RNA levels can be measured to ensure that effective knockdown of the target mRNA was achieved. Briefly, control and knockdown cells are collected and the RNA is harvested, reverse transcribed, and quantified or normalized to an internal control (such as GAPDH). If RNA levels are reduced, then a longer period of time may be needed to achieve a reduction in the protein levels, especially if the protein is present in abundant quantities or has a long half-life. In some cases, when incomplete protein knockdown is observed and mRNA levels are not completely reduced, pooled siRNAs (multiple sequences targeting different segments of the target mRNA) can be introduced into the cells. Detailed descriptions of PCR can be found at Labome.
While RNAi is perhaps the most precise gene silencing mechanism available, it still has both specific and nonspecific off-target effects. The latter is caused by partial sequence complementarity of either the sense or antisense siRNA strands to non-target mRNAs. This is a problem for both siRNAs and shRNAs and is not dependent upon the method of delivery. As little as a 7 nt complementation is capable of producing off-target suppression and is dependent upon the sequence context surrounding the complementary region, the position of the sequence in the mRNA, and the copy number of the sequence within the mRNA.
Due to these specific off-target effects, it is important to test for them through assays such as microarray-based gene expression analysis. siRNAs can be chemically modified to reduce off-target effects. For example, the addition of methyl groups to the 2’ position of the ribosyl ring of the second base of the siRNA can reduce off-target effects. These kinds of chemical modifications are only possible with siRNA oligomers, however. Alternatively, pooled siRNAs against the same gene, as in the form of siPOOL, can reduce the off-target effects [22]. Interestingly, evidence suggests that shRNAs do not have as many off-target effects as siRNAs. While it is not clear why this is the case, it is thought that it may be due to the fact that shRNAs are transcribed in the nucleus, and may be susceptible to further processing. Also, siRNAs may undergo degradation in the cytoplasm, leading to off-target effects.
Non-specific off-target effects involve activation of the interferon and other immune responses to the dsRNA, cellular toxicities caused by the nucleotide construct, and effects resulting from the mode of delivery. Long dsRNAs cause strong induction of the innate immune response, similar to what is observed during viral infection, leading to global mRNA degradation. siRNAs and shRNAs, on the other hand, only induce a partial interferon response. Since some of this activation is sequence dependent, sequence modifications can reduce the immunogenicity of the shRNA or siRNA. As is the case with their specific off-target effects, chemical modification of siRNA oligomers can decrease their ability to induce immune responses. However, these modifications can also reduce their gene silencing ability.
The target specificity that siRNAs and shRNAs provide has made them promising in medical applications as both therapeutic and diagnostic tools. In 2018, FDA approved first siRNA drug, Alnylam Pharmaceuticals’ Onpattro (patisiran), for the treatment of a peripheral nerve disease caused by hereditary transthyretin-mediated amyloidosis, also known as hATTR, in adults. Patisirant is a double-stranded siRNA against both wild-type and mutant transthyretin. They have been used to target oncogenes such as Bcl-2 and p53, as well as k-ras carrying the oncogenic valine-112 mutation. Combinations of siRNAs against multiple targets within cancer cells have also shown promising results. siRNA therapies have also proven effective in mouse models of neurological diseases, such as Huntington’s disease. In addition to genetic disorders, RNAi is being tested in potential therapies for viral infection. Clinical trials are underway for the treatment of macular degeneration, diabetic retinopathy, and hepatitis C. However, there are still obstacles that need to be overcome. As was observed in vitro, there is still a concern that activation of the immune response upon introduction of dsRNA may exhibit excessive toxicity and inhibit effectiveness. Additionally, effective systems to ensure delivery to the target tissues need to be improved upon.
Transfection reagents can be acquired commercially or formulated in the lab. It should be noted that, in addition to the most common transfection reagents that are available (Lipofectamine, fugene, calcium phosphate, protamine sulphate [23], etc.), there are also many commercially formulated kits that are optimized for the transfection of certain cell types (such as Amaxa Human Monocyte Nucleofactor Kit from Lonza [10], the Silencer(R) siRNA Transfection II Kit by Invitrogen/Ambion or specifically for in vivo - for example, in vivo-jetPEI from Polyplus-transfection [24] or Transit-TKO from Takara Bio [25]. Butler AA et al injected Lincode SMARTpool siRNAs conjugated with in vivo-jetPEI into mouse hippocampal areas to evaluate the role of lncRNA Neat1 in memory formation [24].
Commercially available transfection reagents are available. In addition to those discussed below, Batie M et al transfected Hela cells using INTERFERin from Polyplus-transfection with siRNAs purchased from MWG [26]. Wang L et al used the same INTERFERin to transfect RAW264.7 cells [27]. Saito T et al transfected HepG2 cells with SMARTpool siRNAs using Dharmafect 1 from Thermo Fisher [28].
This formulation of the Lipofectamine reagent allows for rapid and consistent transfection of siRNAs into a wide range of cell types, for example, RF/6A cells [25], with high efficiency. It has been used to deliver siUch-L1 from Ambion to HEK-293 cells [29], SMARTpool ON-TARGETplus and Accell siRNA oligonucleotides to human and mouse cells [11, 12], or Qiagen siRNAs to HK2 cells [16]. Lee J et al transfected iPSC-derived cardiomyocytes with either scramble siRNA or siRNA against PDGFRB using it [30]. Yang J et al used Lipofectamine RNAiMAX to transfect a custom siRNA library targeting 2,725 multipass transmembrane proteins to identify the protein-activated chloride channel [17]. Genet G et al transfected FlexiTube and SMARTpool siRNAs into HUVEC cells with RNAiMax [31].
This formulation of Lipofectamine reagent allows for the transfection of siRNAs and shRNA vectors into adherent and suspension cell lines. Lipofectamine 2000 was used to transfect, for example, into HEK293T cells [14, 32] or Hela [32]. Lipofectamine 3000, to A498 cells [32].
Lipofectamine is a cationic lipid with a positively charged head group and 1–2 hydrocarbon chains. The head group interacts with the phosphate backbone of the nucleic acid. The positive surface charge of the liposomes formed by Lipofectamine/RNA mixtures allows for the fusion of liposome/nucleic acid complexes with the negatively charged cell.
Oligofectomine is a proprietary formula that forms stable complexes with nucleic acids to allow for the transfection of siRNAs into eukaryotic cells. It is specific and non-toxic.
The MISSION® siRNA Transfection Reagent allows for the delivery of siRNAs into eukaryotic cells, resulting in >90% silencing efficiency. Using lower concentrations of siRNAs reduces off-target effects.
This system allows for the transfection of siRNAs into hard-to-transfect cell types, including neuronal, primary, differentiated, and non-dividing cells. It is based on a peptide transfection reagent.
H Hu et al use various shRNA lentiviral particles from Santa Cruz Biotechnology to investigate the role of RAK1 in arteriovenous fistula maturation [33]. Freeman SA et al electroporated 100 pM mTPCN1 stealth siRNA from Thermo Fisher (MSS217431) to 2x106 mouse macrophage cells in suspension and achieved a 60-90% reduction in the TPCN1 mRNA level as determined by qPCR [20]. MilliporeSigma offers sets of 3 siRNA duplexes targeting a specific mouse or human gene, with a guarantee that 2 will provide knockdown efficiency of at least 75%. Many companies also offer shRNA expression plasmids as well as shRNA-expressing viral vector systems (Table 6). Leone RD et al ordered TRC lentiviral shRNAs targeting a number of genes from MilliporeSigma [34]. Zhou R et al obtained the adenovirus expressing CHC shRNA from SignaGen Laboratories and those expressing TrkB, FGFR1 or calpain-2 shRNA from Vector BioLabs [35]. Kitchen P et al used pGFP-C-shLenti-AQP4 and pGFP-C-shLenti-Control from OriGene to infect dorsal column neurons in rats [36]. Deming et al obtained MS4A4A shRNA pGFP-C-shLenti (TL303135B) and the control hRNA pGFP-C-shLenti Scrambled (TR30021) from OriGene to transduce cultured monocyte-derived macrophages [37]. Chakraborty AA et al obtained pLKO-based shRNA plasmids targeting Kdm6a and Kdm6b from GPP, Broad Institute [38]. Lee YR et al used MilliporeSigma siRNA duplexes and Dharmacon lentivirus-based shRNA constructs to target WWP1 gene and Dharmacon SMARTpool siRNA to target MYC [39]. Huang H et al purchased from Dharmacon shRNA lentiviral vectors against SETD2, METTL3, METTL14, WTAP, Mettl14 and the control (shCtrl) to knockdown their expressions in mouse embryonic stem cells, HeLa and/or HepG2 cells [32]. Moro A et al knocked down DROSHA expression in HUVEC with Dharmacon shRNA SMARTvectors from GE Healthcare [40].
| Company | Use | Backbone-Promoter | Selectable Marker | Reference |
|---|---|---|---|---|
| MilliporeSigma | shRNA expression | pLKO.1-CMV PLKO.1-UbC | eGFP, tGFP, TagCFP, TagYFP, TagRFP, TagFP635, TurboGFP, and TagFP635 Puromycin, neomycin | |
| Inducible shRNA expression | pLKO-puro-IPTG-1xLacO | Puromycin | ||
| pLKO-puro-IPTG-3xLacO | Puromycin | |||
| Transient or Stable shRNA transfection and lentivirus production | TRC2-pLKO | Puromycin | ||
| SBI System Biosciences | Backbone/promoter for shRNA | pSIH1-H1 pSIF-HI pGreenPuro pFIV-H1 | GFP, RFP, Puromycin, Hygromycin, Neomycin and Zeocin | |
| Backbone/promoter for microRNA | pCDH-CMV-MCS-EF1 pMIF-cGFP-Zeo | GFP, RFP, Puromycin, Hygromycin, Neomycin and Zeocin | ||
| Addgene | Retroviral vector for shRNA expression | pMKO.1 and pMKO.1 | Puromycin, neomycin, Zeocin, GFP | |
| Lentiviral shRNA expression | pLKO.1 | Puromycin, Hygromycin, neomycin, blasticidin | [41, 42] | |
| Tet-inducible lentiviral shRNA expression | Tet-pLKO | |||
| Conditional shRNA expression under the Cre-Lox control. | pSico | GFP (though cassette can be removed and replaced easily) |
Addgene is a nonprofit repository for plasmids and offers a wide variety of lentiviral and retroviral transfer vectors [43, 44]. It also provides many ready-made shRNA plasmids such as pLKO-RB1-shRNA63 and pLKO-RB1-shRNA19 from T. Waldman (Addgene plasmids 25641 and 25640) [45]. The popular AddGene shRNA ready-made plasmids include GFP shRNA [46], Rictor_1 shRNA [47], beta-catenin shRNA [48], and Raptor_1 shRNA [49].
- Cecconi F, D'Amelio M. Apoptosome. Lingor, P. Regulation of Cell Death and Survival by RNA Interference – The Roles of miRNA and siRNA. Netherlands: Springer 2010. 95-117p. Available from: link.springer.com/chapter/10.1007%2F978-90-481-3415-1_5/fulltext.html
- Choosing Your Perfect Plasmid Backbone. Available from: www.addgene.org/empty_backbones/
- Lentiviral Plasmids. Available from: www.addgene.org/lentiviral/
- Materials and Methods [ISSN : 2329-5139] is a unique online journal with regularly updated review articles on laboratory materials and methods. If you are interested in contributing a manuscript or suggesting a topic, please leave us feedback.
- method