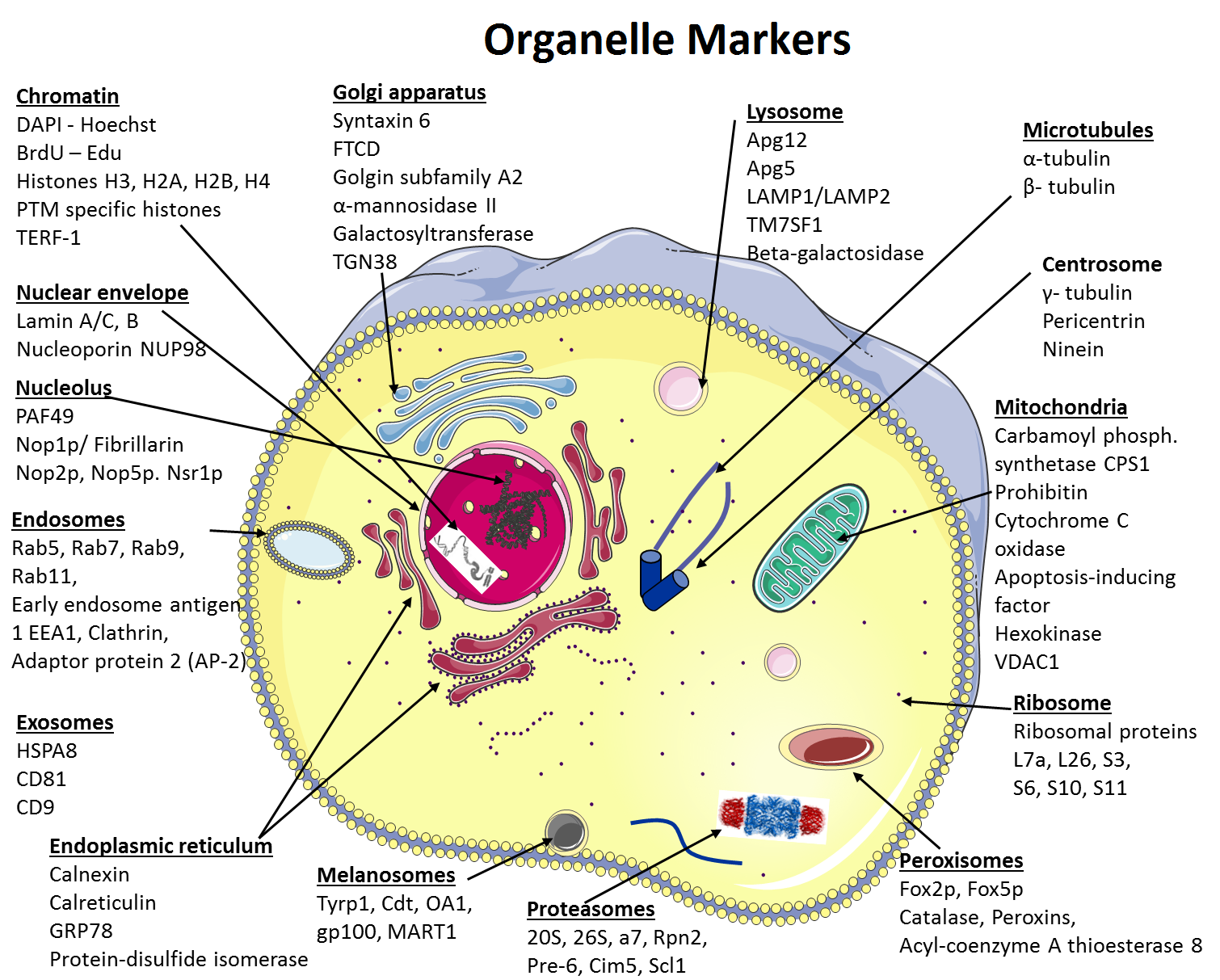
A compilation of organelle markers and the most cited antibodies against these markers among the over 60,000 formal publications Labome has curated for Validated Antibody Database.
Cells are not just sacs of protoplasm. They contain several organelles and structures, each one of them with specific functions. For detailed characterization of an organelle and its composition, purification is most of the times a prerequisite. In addition, characterization of a novel protein requires knowledge of its localization. Using as references other proteins whose localization is well established (markers) is a useful strategy. In this review, these proteins used as markers of organelles in immunoblotting and immunofluorescence are discussed (Figure 1 and Table 1).
| Protein | Full Name | Top three suppliers | Ref | ||
|---|---|---|---|---|---|
| Golgi | |||||
| AP1G1 / gamma-adaptin | adaptor related protein complex 1 subunit gamma 1 | Santa Cruz Biotechnology sc-398867 (1) | [5] | ||
| giantin | Golgin subfamily B member 1 | Abcam ab37266 (13), Enzo Life Sciences ALX-804-600-C100 (7) | [6] | ||
| GM130 | Golgin subfamily A member 2 | BD Biosciences 610822 (146), Abcam ab52649 (32), Cell Signaling Technology 12480 (11) | [5] | ||
| GORASP1 / GRASP65 | golgi reassembly stacking protein 1 | Santa Cruz Biotechnology sc-365434 (1) | [7] | ||
| TGN38/46 | trans-golgi network protein 2 | Invitrogen MA3-063 (10), Santa Cruz Biotechnology sc-271624 (1), MilliporeSigma SAB4200235 (1) | [5, 8] | ||
| Mitochondria | |||||
| AIF | apoptosis-inducing factor | Santa Cruz Biotechnology sc-13116 (16), Cell Signaling Technology 5318 (15), Invitrogen MA5-15880 (3) | |||
| COX | cytochrome c oxidase subunit 1 | Abcam ab14705 (69), Invitrogen 459600 (67) | |||
| VDAC1 | outer mitochondrial membrane protein porin 1 | Abcam ab14734 (82), Cell Signaling Technology 4661 (40), Santa Cruz Biotechnology sc-390996 (14) | |||
| CPS1 | carbamoyl-phosphate synthase | Abcam ab129076 (2) | |||
| PHB | prohibitin | Invitrogen MA5-12858 (70), Abcam ab75766 (4), Santa Cruz Biotechnology sc-377037 (2) | |||
| HK1 | hexokinase-1 | Cell Signaling Technology 2024 (25), Santa Cruz Biotechnology sc-46695 (2), MilliporeSigma WH0003098M1 (2) | |||
| Chromatin | |||||
| HIST1H3F | histone H3.1 | Cell Signaling Technology 4499 (112), Abcam ab10799 (18), MilliporeSigma H9908 (16) | |||
| TERF1 | telomeric repeat-binding factor | Abcam ab10579 (10), Santa Cruz Biotechnology sc-56807 (3), Invitrogen MA1-46089 (2) | |||
| HIST1H4A | histone H4 | Cell Signaling Technology 2935 (7), Invitrogen MA5-14816 (1), Abcam ab197515 (1) | |||
| TERF2IP | telomeric repeat-binding factor 2-interacting protein 1 | Santa Cruz Biotechnology sc-53434 (1), Abcam ab14404 (1) | |||
| HIST1H2BB | histone H2B | Cell Signaling Technology 5546 (11) | |||
| CBX5 | heterochromatin protein 1alpha | Abcam ab109028 (3), Cell Signaling Technology 2623 (2) | |||
| CBX1 | heterochromatin protein 1beta | Abcam ab10811 (5), Cell Signaling Technology 8676 (3) | |||
| CBX3 | heterochromatin protein 1gamma | Abcam ab217999 (2), Santa Cruz Biotechnology sc-398562 (1) | |||
| CENPA | centromere protein A | Enzo Life Sciences ADI-KAM-CC006-E (3), MBL International D115-3 (1) | |||
| CENPC | centromere protein A | Abcam ab50974 (3) | |||
| Nuclear envelope | |||||
| LMNA | prelamin-A/C | Cell Signaling Technology 4777 (50), Santa Cruz Biotechnology sc-7292 (49), Abcam ab8984 (14) | |||
| LMNB | lamin B1 | Invitrogen 33-2000 (37), Cell Signaling Technology 12586 (16), Santa Cruz Biotechnology sc-6217 (12) | |||
| NUP98 | nuclear pore complex protein Nup98 | Cell Signaling Technology 2598 (16), Abcam ab50610 (5), BioLegend 406108 (1) | |||
| EMD | emerin | Leica Biosystems NCL-EMERIN (6), Santa Cruz Biotechnology sc-25284 (3), Abcam ab156871 (2) | |||
| Nucleoplasm | |||||
| KDM1/LSD1 | lysine demethylase 1A | Cell Signaling Technology 2184 (14), Abcam ab129195 (4), Santa Cruz Biotechnology sc-271720 (1) | [9] | ||
| Nucleolus | |||||
| FBL | fibrillarin | Cell Signaling Technology 2639 (24), Abcam ab4566 (20), Novus Biologicals NB300-269 (8) | |||
| FUS | used in Sarcoma | Santa Cruz Biotechnology sc-47711 (30), BD Biosciences 611385 (3) | |||
| Endosomes | |||||
| AP2S1 | AP-2 complex subunit sigma | Abcam ab128950 (3) | |||
| CLTB | clathrin light chain B | Santa Cruz Biotechnology sc-12735 (4), Invitrogen MA5-11860 (1), MilliporeSigma C1985 (1) | |||
| CLTC | clathrin heavy chain 1 | Invitrogen MA1-065 (66), BD Biosciences 610499 (31), Cell Signaling Technology 4796 (8) | |||
| EEA1 | early endosome antigen 1 | BD Biosciences 610457 (66), Cell Signaling Technology 3288 (56), Abcam ab70521 (16) | [7, 10] | ||
| Rab5 | Ras-related protein Rab5 | Cell Signaling Technology 3547 (45), Santa Cruz Biotechnology sc-46692 (14), BD Biosciences 610281 (6) | [9] | ||
| Rab7 | Ras-related protein Rab-7a | Cell Signaling Technology 9367 (70), Abcam ab50533 (25), Enzo Life Sciences ALX-804-961-0100 (10) | [9] | ||
| Rab9 | Ras-related protein Rab-9a | Invitrogen MA3-067 (12), Cell Signaling Technology 5118 (11), Abcam ab2810 (2) | |||
| Rab11 | Ras-related protein Rab-11A | Cell Signaling Technology 5589 (29), BD Biosciences 610656 (16), Invitrogen 700184 (5) | |||
| TFRC | transferrin receptor | Invitrogen 13-6800 (454), BD Biosciences 555534 (18), BioLegend 334102 (10) | [7] | ||
| Endoplasmic reticulum (KDEL, a motif, is not listed here) | |||||
| CANX | calnexin | Cell Signaling Technology 2679 (40), Invitrogen MA3-027 (16), BD Biosciences 610523 (13) | [7] | ||
| CALR | calreticulin | Cell Signaling Technology 12238 (19), Abcam ab92516 (10), BD Biosciences 612136 (9) | |||
| GRP78 | 78 kDa glucose-regulated protein | Cell Signaling Technology 3177 (108), BD Biosciences 610978 (29), Santa Cruz Biotechnology sc-376768 (9) | |||
| PDI | Protein disulfide-isomerase (PDI) | Invitrogen MA3-019 (46), Cell Signaling Technology 3501 (34), Enzo Life Sciences SPA-890 (5) | |||
| SEC13 | SEC13 homolog, nuclear pore and COPII coat complex component | Santa Cruz Biotechnology sc-514308 (2) | [11] | ||
| SEC61B | SEC61 translocon subunit beta | Santa Cruz Biotechnology sc-393633 (2) | [9] | ||
| Microtubules | |||||
| TUBA1A | alpha-tubulin | MilliporeSigma T9026 (423), Invitrogen 62204 (113), Santa Cruz Biotechnology sc-8035 (90) | |||
| TUBB3 | beta tubulin III | BioLegend 801201 (255), MilliporeSigma T8660 (89), Invitrogen 32-2600 (36) | [10] | ||
| Centrosome | |||||
| TUBG1 | gamma-tubulin | MilliporeSigma T6557 (166), Santa Cruz Biotechnology sc-17787 (12), Invitrogen MA1-850 (10) | [8] | ||
| Actin filaments | |||||
| ACTA2 | actin alpha (smooth muscle) | MilliporeSigma A2547 (333), Invitrogen MA5-11869 (290), Abcam ab7817 (125) | |||
| Autophagosomes and lysosomes | |||||
| ATG12 | autophagy-related protein 12 | Cell Signaling Technology 4180 (28), Santa Cruz Biotechnology sc-271688 (2), GeneTex GTX629815 (1) | |||
| ATG5 | autophagy protein 5 | Cell Signaling Technology 12994 (52), Abcam ab108327 (21), Santa Cruz Biotechnology sc-133158 (10) | |||
| CD68 | CD68 | Dako M0876 (92), Invitrogen MA5-13324 (91), Abcam ab955 (54) | [12] | ||
| GLB1 | beta-galactosidase | LifeSpan Biosciences LS-B10217 (2), Santa Cruz Biotechnology sc-377257 (1) | |||
| LAMP1 | lysosome-associated membrane protein 1 | Cell Signaling Technology 9091 (58), BD Biosciences 555798 (58), Santa Cruz Biotechnology sc-20011 (56) | [10, 13] | ||
| LAMP2 | lysosome-associated membrane protein 2 | Santa Cruz Biotechnology sc-18822 (47), Developmental Studies Hybridoma Bank H4B4 (34), Abcam ab25631 (32) | [9, 14] | ||
| LC3 | microtubule-associated proteins 1A/1B light chain 3A | Cell Signaling Technology 12741 (91), MBL International M152-3 (36), Nanotools 0260-100/LC3-2G6 (9) | [10] | ||
| Melanosomes | |||||
| OA1 | melanoma antigen recognized by T-cells 1 | Dako M7196 (22), Invitrogen MA5-15237 (19), Santa Cruz Biotechnology sc-20032 (6) | |||
| PMEL | melanocyte protein PMEL | Invitrogen MA5-13232 (62), Abcam ab137078 (6) | |||
| TYRP1 | tyrosinase-related protein 1 | Abcam ab3312 (5), BioLegend 917801 (3), Invitrogen MA5-12293 (2) | |||
| TYRP2 | tyrosinase-related protein 2 | Santa Cruz Biotechnology sc-74439 (4) | |||
| Peroxisomes | |||||
| CAT | catalase | Cell Signaling Technology 14097 (7), Santa Cruz Biotechnology sc-271803 (3), Abcam ab76024 (2) | |||
| Ribosomes | |||||
| RPS6 | ribosomal protein s6 | Cell Signaling Technology 2217 (236), Santa Cruz Biotechnology sc-74459 (15), Invitrogen 14-9007-37 (1) | |||
| Proteasomes | |||||
| PSMA1 | 20S proteasome alpha-subunit | Enzo Life Sciences BML-PW8195-0100 (5), Abcam ab22674 (3), Santa Cruz Biotechnology sc-58412 (1) | |||
| PSMD1 | 26S proteasome non-ATPase regulatory subunit 1 | Santa Cruz Biotechnology sc-166038 (2), Enzo Life Sciences BML-PW9270-0025 (1) | |||
| PSMD7 | 26S protease regulatory subunit 7 | Santa Cruz Biotechnology sc-390705 (1) | |||
| Mitosis | |||||
| HIST1H3F | phosphohistone H3 | Cell Signaling Technology 4499 (112), Abcam ab10799 (18), MilliporeSigma H9908 (16) | |||
| Cytokinesis | |||||
| 9212 | Aurora B kinase | Cell Signaling Technology 2914 (27), BD Biosciences 611082 (15), Abcam ab3609 (7) | |||
The main function for Golgi apparatus is the proper folding of macromolecules and their secretion to the extracellular environment (exocytosis via the trans-Golgi network), together with other proteins and lipids, in the intracellular environment (Golgi stack). Golgi can also synthesize proteoglycans and carbohydrates. They are usually disassembled during mitosis and reassembled again after mitosis in each daughter cell [15].
The receptor binding cancer antigen expressed on SiSo cells is a type III transmembrane Golgi protein [16] and more specifically localized at ER-Golgi intermediate compartment and the cis-Golgi [17].
Syntaxin 6 is mainly found in Golgi and is involved in trafficking of intracellular vesicles [18].
FTCD is a 58 kDa enzyme with transferase and deaminase activity. It is localized on Golgi and facilitates bundling of vimentin starting from Golgi, but recently has also been found in the centrosome [19].
GM130 is a Golgi auto-antigen which probably has a function in ER-Golgi transport [20]. Another golgin member, giantin, also serves as a Goglgi marker. Deming et al colocalized TREM2 with Golgi in human macrophages using giantin as a marker [6].
Alpha-mannosidase II is localized on the membrane of the Golgi apparatus and is involved in protein glycosylation, as it regulates steps of N-glycan synthesis.
B4Gal-T6 is involved in the biosynthesis of glucosphingolipids and is another Golgi internal membrane marker.
TGN38 regulates membrane traffic from the trans-Golgi network (the secretory mechanism) to the plasma membrane. Upon Brefeldin A treatment, the Golgi stack is de-organized and the trans-Golgi network collapses upon the centrosome. Thus, TGN38 staining distinguishes the TGN from the Golgi stack.
The mitochondrion (greek mitos, μίτος = thread + chondrion, χονδρίον = granule) is an organelle of 0.5-1.0 μm in diameter. They are considered as the cellular power plants because they synthesize energy in the form of Adenosine Triphosphate (ATP) but they also have other functions. The mitochondrion is composed of the inner and outer membranes, the inter-membrane space, the cristae and the matrix while they contain their own DNA separated from the nuclear. In humans, more than 600 distinct proteins have been found [21] and some of them are used as markers. Mitochondria can also be easily identified through dyes such as MitoTracker.
In addition to the markers discussed below, Ling Q et al used Slp1 as a mitochondrial marker in Arabidopsis protoplasts [22]. TOM20, an outer mitochondrial membrane protein serves as a mitochondrial marker in brain neurons [9, 13].

is the 163 kDa mitochondrial isozyme of this enzyme, which is involved in the urea cycle and removes excess of ammonia from the cell. CPS1 is a marker of liver and kidney mitochondria.
is a 30 kDa protein of the inner mitochondrial membrane and probably regulates mitochondrial respiration. It is involved in several activities including apoptosis, cell cycle regulation and senescence. Prohibitin is most abundant during the G1 phase of the cell cycle and upon treatment with thiamphenicol, a mitochondrial protein synthesis inhibitor [23].
is protein complex of the inner mitochondrial membrane [24]. It is involved in the translocation of protons and catalysis of oxygen to water required for ATP synthesis. Most of the COX subunits can be good mitochondrial markers.
is a 67 kDa protein of the inter-membrane space and is ubiquitously expressed. While in mitochondria, AIF functions as an oxidoreductase and has anti-apoptotic activity. However, during apoptotic signals, AIF is released to the cytoplasm and translocates to the nucleus to induce nuclear apoptosis.
is a 100 kDa kinase of the outer mitochondrial membrane catalyzing the first step of glycolysis. Hexokinase phosphorylates hexoses (a six-carbon sugar) to form hexose phosphates (e.g., glucose to glucose-6-phosphate).
is a 100 kDa kinase of the outer mitochondrial membrane catalyzing the first step of glycolysis. Hexokinase phosphorylates hexoses (a six-carbon sugar) to form hexose phosphates (e.g., glucose to glucose-6-phosphate).
(outer mitochondrial membrane protein porin 1) is the outer mitochondrial membrane receptor for hexokinase and BCL2L1.
Mitochondrial proteins binding DNA and RNA, including TFAM, POLG, ERAL1, LRPPRC and p32 were shown to be involved in the regulation of mitochondrial homeostasis. One of these proteins, TFAM, regulates mtDNA transcription. With regard to the clinical significance, inhibition of TFAm-mediated DNA transcription may lead to the development of Parkinson’s and Alzheimer’s diseases [25].
In addition, POLG (mitochondrial DNA polymerase γ) is involved in the mechanisms of mitochondrial replication. Clinical data showed that mutations in the POLG gene were associated with progressive ophthalmoplegia [26]. Also, transgenic mice carrying a POLG gene mutation were shown to develop severe cardiomegaly [27].
Another mitochondrial protein, ERAL1, is localized in the mitochondrial matrix and downregulation of its expression may cause mitochondrial disorders and growth abnormalities [28]. With regard to LRPPRC, mutations in the LRPPRC gene were detected in patients with Leigh syndrome [29]. Also, mutations in the gene encoding RNA binding protein p32 are associated with the mitochondrial respiratory disorder [30].
There are several proteins used to distinguish the distinct nuclear structures from each other.
The DNA molecule is condensed as it is wrapped around the four core histones (H2A, H2B, H3, H4 - which form an octamer) and form the nucleosome. Histone H1 is a protein linker which binds to distant chromatin areas and compacts it further. The DNA with its histones and other proteins that associated with it (to regulate transcription, replication, DNA repair, etc.) is called chromatin.
Therefore, any dye that binds DNA can be used as a chromatin marker. For example, 4',6-diamidino-2-phenylindole (DAPI) [31] and the dyes Hoechst 33258 and Hoechst 33342 are the most common. Bromodeoxyuridine (BrdU) is a synthetic nucleoside used to detect proliferating cells. 5-ethynyl-2'-deoxyuridine (EdU) staining is equally effective.
All core histones are required for nucleosome formation. Therefore, anti-histone antibodies would mark mainly chromatin. Histone modification-specific antibodies can distinguish between euchromatin (e.g., H3K4me3, H3K4me1, H3K9ac, H3K27ac or H3K36me3) and heterochromatin (e.g., H3K9me3, H3K27me3, H4K20me3). Phosphohistone H3S10ph can also be used to indicate the mitotic state of cells [32]. Heterochromatin protein 1 (HP1) binds to histone tails, and more specifically to trimethylated lysine of H3 (H3K9me3) and contributes to closed chromatin state, therefore is a great marker of heterochromatin. Ling Q et al used anti-histone H3 to identify nuclei in Arabidopsis protoplasts [22].
Telomeric repeat-binding factor 2-interacting protein 1 (TERF-1, RAP1) is localized at telomeres and regulates telomere length.
CENP-A is a histone H3 variant that localizes at centromeres, the central, condensed structure of chromosomes to which the spindle is attached during chromosome segregation. Alternatively, CENPC can be used.
The nuclear envelope is the lipid bilayer membranous structure surrounding the nucleus and separates it from the cytoplasm. The inner membrane is comprised of a network of intermediate filaments, the lamina, while the outer membrane is physically linked to the endoplasmic reticulum, thus sharing some common proteins. Small holes on the nuclear envelope constitute the nuclear pore complexes of about 100 nm diameter and they connect the inner with the outer nuclear membrane as well as import or export proteins to and from the nucleus.
Lamin A (74 kDa) and Lamin C as well as Lamin B (68 kDa) can be detected by antibodies to show the nuclear envelope [9], which is disassembled during mitosis. During apoptosis, lamin A and C will be cleaved into two fragments of 40-50 kDa and of 28 kDa.
Nucleoporin 98 (NUP98) belongs to the nuclear pore complex. Emerin is another inner nuclear membrane marker.
The nucleolus is a non-membranous structure inside the nucleus of the cell which transcribes and assembles ribosomal RNAs. The protein components of the nucleoli can be used as markers, such as the RNA polymerase PAF49, the nucleolar protein 1 (Nop1p/Fibrillarin), Nop2p, Nop5p and Nsr1p.
FUS (Fused in Sarcoma) protein staining normally stains the nucleus without its nucleoli.
Endosomes are cytoplasmic compartments which encircle molecules and transfer them from the membrane to other parts in the cell. Usually, endocytosed complexes (e.g., receptor-ligand) are separated in early endosomes and each component can be transferred to its new destination (e.g., lysosomes, Golgi, etc.) via the late endosomes. LysoTrackers (green or red) are popular dyes to label lysosome [33, 34]. Flaherty SE et al stained lysosome in bone marrow-derived macrophages with LysoTracker from Invitrogen [33].
Lysobisphosphatidic acid is a phospholipid exclusively located in late endosomes and regulates endosomal cholesterol levels [35]. It is used as a late endosome marker [14], and detected through antibodies.
Rab5 is a small GTPase (24 kDa) of the Ras family which shuttles from the plasma membrane to early endosomes and regulates vesicular trafficking and fusion of plasma membranes with early endosomes, via its interaction with other proteins. Similarly, Rab7 [14, 36], Rab9, and Rab11 [14] are equally good endosomal protein markers. Laflamme C et al used GFP-Rab7a and GFP-Rab9 as the markers for vesicle trafficking to lysosomes, GFP-Rab5a and GFP-Rab11a as markers of early and recycling endosomes, respectively [37].
One of the Rab5-interacting proteins is the early endosome antigen 1 (EEA1). It is a 162 kDa protein and participates in endosomal trafficking. EEA1 is one of most commonly used markers for early endosome, for example, during a study of the interaction between segmented filamentous bacteria and intestinal epithelial cells [14] or the role of ABCA7 in microglia [36] or the resorption of water and others from macropinosomes [38].
Other structures for endocytosis and transfer of molecules are the clathrin-coated pits (or vesicles). These vesicles consist of a proteinaceous coat which packs membrane receptors and other molecules. Clathrin and Adaptor protein-2 (AP-2) are excellent markers of clathrin-coated vesicles. However, some endosomes arise through clathrin-independent pathway [14], thus, clathrin, and AP-2 do not label all endosomes.
Sometimes endosomes fuse with the plasma membrane and are secreted into the extracellular environment and they constitute a secretion mechanism. Exosomes are important as they contain cytoplasmic and membrane proteins as well as lipids and RNA molecules that might be potential biomarkers of particular diseases. For more details, please see Labome's review dedicated to exosomes.
The endoplasmic reticulum (ER) is a cytoplasmic structure containing many chaperones that help polypeptides to fold properly and to assemble protein complexes. Most ER proteins contain the KDEL motif (Lysine-Aspartate-Glutamine-Leucine) and are retained through interaction with an internal ER KDEL receptor. Therefore, an anti-KDEL antibody recognizing the motif is used as an ER-positive marker. For example, Deming et al colocalized TREM2 with endoplasmic reticulum in human macrophages using the KDEL marker [6]. Other markers include SEC61B [9] and SEC13 [11].
Calnexin is a 90 kDa integral ER membrane protein which binds unfolded proteins and retains them at ER.
Calreticulin is 48 kDa chaperone of the ER with a KDEL motif at the C-term and binds monoglucosylated proteins synthesized in the ER. Ling Q et al marked ER in Arabidopsis protoplasts with an anti-calreticulin antibody [22].
The 78 kDa glucose-regulated protein (GRP 78) contains a KDEL C-term motif and facilitates the assembly of protein complexes in the ER. It is essential for cell viability.
PDI habitat is the ER due to its KDEL domain. PDI has several functions, including the formation of disulfide (S-S) bonds on unfolded proteins.
Microtubules are elongated filaments consisting of tubulins. Alpha-tubulin and β-tubulin are globular proteins of 55 kDa which form heterodimers and they polymerize to form the cylindrical microtubule. They are involved in numerous cellular functions and especially cell structure maintenance, intracellular transport, or the formation of mitotic spindles that separate the sister chromatids during cell division. Tubulin polymerization starts at the centrosome, which constitutes the microtubule organizing center (MTOC) in interphase and the spindle poles during mitosis, where distinct protein complexes constitute the scaffold for tubulin polymerization initiation. These complexes contain γ-tubulin. The centrosome has only one copy per cell, which will duplicate during mitosis.
Antibodies targeting α- or β-tubulin are good markers for microtubules. Tornabene P et al, for example, used MilliporeSigma T6793, which recognizes acetylated tubulin, to label microtubules in Hela cells [39].
Anti-γ-tubulin antibodies show the centrosome [8]. Other centrosomal markers are the pericentrin and ninein [8]. Pericentrin is a 220 kDa protein involved in the initial formation (nucleation) of the microtubule. Ninein is a centrosomal protein involved in microtubule nucleation and capping of the minus- and plus-ends.
Similarly to microtubules, actin filaments are double helical thin cylindrical tubes made of α- or β-actin. Their dynamic polymerization and depolymerization cycles regulate cell movement, cell polarization and scaffolding of the cell. In addition, several actin-binding proteins control actin polymerization.
They can be labeled by anti-actin antibodies that bind to actin monomers or the fluorescently labeled toxin phalloidin that binds to filamentous actin, for example, Alexa Fluor 647 phalloidin from Invitrogen [40].
Recently, SIR-actin and SIR tubulin, cell permeable probes has become commercially available for live cell imaging [8, 41]. Similarly, the actin chromobody can be used to study live actin dynamics upon plasmid transfections in living cells. LifeAct peptide can also be used to label F-actin [42].
Autophagosomes are intracellular organelles formed by elongation of small membrane structures, the autophagosome precursors, into a double membrane which surrounds damaged organelles. The autophagosome would then fuse with a lysosome, which contains hydrolases and other degradation enzymes, and is set for degradation [43]. Please see Labome dedicated review on autophagy.
Apg12 and Apg5 are covalently associated with each other and function as a unit. Therefore, the Apg12-Apg5 conjugate is localized at the autophagosome membrane during elongation. Therefore it is a good marker for the initiation of autophagy. Several other Apg isoforms regulate autophagosome formation and its fusion to the lysosome and they can be used as autophagosomal markers.
The microtubule-associated protein 1 light chain 3 (LC3) is also localized at the membrane but when fully formed it is found at the isolation membranes as well. LC3 can be found on lysosomes but with less abundance [43]. Caution has been raised as to whether Western blot of LC3 and its associated protein sequestosome 1 (SQSTM1, also known as p62) [44] can truly reflect the status of autophagosomes, especially in the case of p62 [45].
Lysosome Associated Membrane Protein 1 and 2 (LAMP1 [13, 36] ; LAMP2 [9, 14] ) are components of the lysosomal membrane and therefore constitute excellent lysosomal markers. Aikawa T et al detected lysosomes in mouse brain sections and primary microglia culture with Abcam's ab25630 anti-LAMP1 antibody [36]. Laflamme C et al used LAMP1 as a marker of lysosomes to localize C9ORF72 [37].
Beta-galactosidase is one of the glycosidases that can be used as lysosomal markers. Proteases such as cathepsin S (CTSS) and D (CTSD) have been used as lysosomal markers as well [9, 44].
Cation-dependent mannose-6-phosphate receptor (M6PR) is involved in the transport of lysosomal enzymes from the Golgi and cell surface to the lysosomes.
A full list of about 77 proteins which are primarily found in the lysosome can be found in Lübke et al [46], although not all of them are established as markers.
Melanosomes are organelles of melanocytes, skin and retinal epithelial cells. Melanosomes contain the pigment melanin which protects cells from harmful ultraviolet (UV) radiation. Markers for melanosomes can be tyrosinase, Tyrp1, Dct, OA1, gp100, and MART1 [47].
Peroxisomes are structures which house oxidative reactions, such as fatty acid β-oxidation, and protect from peroxides [48].
Fox2p and Fox5p are peroxisomal membrane receptors.
Catalase is a peroxisomal protein which protects cells from the toxic effects of hydrogen peroxide. It is expressed in the small intestine [49].
Acyl-coenzyme A thioesterase 8 belongs to a group of enzymes that catalyze the hydrolysis of acyl-CoAs to the free fatty acid and coenzyme A (CoASH), providing the potential to regulate intracellular levels of acyl-CoAs, free fatty acids and CoASH.
Peroxins (a class of 24 genes) are integral parts of peroxisome development. Ling Q et al identified peroxisome from Arabidopsis protoplast lysates with anti-PEX13 and anti-PEX14 antibodies [22].
The structures at the cellular membranes that provide attachment to the extracellular matrix and cell-to-cell adhesion and communication are called intercellular junctions and are of three different types: tight, gap and adherent junctions. Cadherins and integrins are integral components of all junctions and therefore are excellent markers, for example, adherens junctions marker E-cadherin, apical marker ZO-1 [50], and basolateral marker alpha6 integrin [42]. Connexin 43 (or gap junction protein alpha 1 – GJA1) is characteristic of gap junctions [51].
Ribosomes are molecular complexes of proteins and RNA molecules (ribonucleoprotein) in which proteins are synthesized. They are comprised of a small 40S subunit and a large 60S subunit. Several ribosome-specific proteins can be used as markers.
Antibodies against ribosomal proteins L7a, L26 (a component of the 60S subunit), S3, S6, S10, S11 (40S subunit) are characteristic examples.
Due to their importance for cell proliferation and development, ribosomal proteins (RP) may be selected as potential clinical diagnostic markers. In particular, ribosomal disorders, which are usually characterized by the development of anemia and growth abnormalities, induce specific changes in the expression of ribosomal proteins. One of these disorders, Diamond Blackfan Anemia (DBA), is characterized by the mutations in genes encoding S19 [52], S24 [53], S17 [54] and L5 [55] ribosomal proteins. Also, the development Shwachman Diamond syndrome (SDS) characterized by bone marrow and skeletal abnormalities, is accompanied by the downregulation of the expression of several RPs, such as S9, S20, L6, and L15 [56].
In addition, RPs were shown to be affected in cardiovascular and metabolic pathologies. In particular, the knockdown of S15a negatively affects heart function in Drosophila [57]. Another RP, S6K1, was shown to be involved in the development of type 2 diabetes [58].
Proteasomes are multi-protein complexes and their function is to degrade proteins by proteolysis. Each proteasome consists of four stacked rings forming a central pore, the core. A seven-protein complex (β subunits) of proteolytic enzymes forms the two rings in the interior while seven α subunits form the entrance through which proteins enter and reach the core.
The proteasome subunits 20S proteasome [59], 26S proteasome, α7 and Rpn2 [60], Pre6, Cim5 and Scl1 [61] are commonly used as proteasomal markers.
Histone H3 is phosphorylated at Serine 10 (H3S10ph) and is essential for the onset of mitosis [32].
Aurora B kinase [32].
Primary cilia are membrane protrusions of epithelial cells that form upon specific conditions. Microtubules not only support the cilium structurally but provide a path for proteins to translocate to this organelle. Such proteins can be used as cilia markers and are Inversin, Polaris, Arl13b [7] and others [62].
- Granell S, Baldini G, Mohammad S, Nicolin V, Narducci P, Storrie B, et al. Sequestration of mutated alpha1-antitrypsin into inclusion bodies is a cell-protective mechanism to maintain endoplasmic reticulum function. Mol Biol Cell. 2008;19:572-86 pubmed
- Lipp J, Hirota T, Poser I, Peters J. Aurora B controls the association of condensin I but not condensin II with mitotic chromosomes. J Cell Sci. 2007;120:1245-55 pubmed
- Nakajima K, Ono K, Nishikawa S, Hieda W, Yoshida T. Interconversion of molecular size of DNA polymerase from Rauscher leukemia virus. Bibl Haematol. 1975;:603-5 pubmed
- Bock J, Lin R, Scheller R. A new syntaxin family member implicated in targeting of intracellular transport vesicles. J Biol Chem. 1996;271:17961-5 pubmed
- Hagiwara H, Tajika Y, Matsuzaki T, Suzuki T, Aoki T, Takata K. Localization of Golgi 58K protein (formiminotransferase cyclodeaminase) to the centrosome. Histochem Cell Biol. 2006;126:251-9 pubmed
- Weide T, Bayer M, Koster M, Siebrasse J, Peters R, Barnekow A. The Golgi matrix protein GM130: a specific interacting partner of the small GTPase rab1b. EMBO Rep. 2001;2:336-41 pubmed
- Taylor S, Fahy E, Zhang B, Glenn G, Warnock D, Wiley S, et al. Characterization of the human heart mitochondrial proteome. Nat Biotechnol. 2003;21:281-6 pubmed
- Coates P, Nenutil R, McGregor A, Picksley S, Crouch D, Hall P, et al. Mammalian prohibitin proteins respond to mitochondrial stress and decrease during cellular senescence. Exp Cell Res. 2001;265:262-73 pubmed
- Ostermeier C, Iwata S, Michel H. Cytochrome c oxidase. Curr Opin Struct Biol. 1996;6:460-6 pubmed
- Bertram L, Hsiao M, Lange C, Blacker D, Tanzi R. Single-nucleotide polymorphism rs498055 on chromosome 10q24 is not associated with Alzheimer disease in two independent family samples. Am J Hum Genet. 2006;79:180-3; author reply 183-4 pubmed
- Longley M, Clark S, Yu Wai Man C, Hudson G, Durham S, Taylor R, et al. Mutant POLG2 disrupts DNA polymerase gamma subunits and causes progressive external ophthalmoplegia. Am J Hum Genet. 2006;78:1026-34 pubmed
- Lewis W, Day B, Kohler J, Hosseini S, Chan S, Green E, et al. Decreased mtDNA, oxidative stress, cardiomyopathy, and death from transgenic cardiac targeted human mutant polymerase gamma. Lab Invest. 2007;87:326-35 pubmed
- Mizushima N, Ohsumi Y, Yoshimori T. Autophagosome formation in mammalian cells. Cell Struct Funct. 2002;27:421-9 pubmed
- Basrur V, Yang F, Kushimoto T, Higashimoto Y, Yasumoto K, Valencia J, et al. Proteomic analysis of early melanosomes: identification of novel melanosomal proteins. J Proteome Res. 2003;2:69-79 pubmed
- Purdue P, Lazarow P. Peroxisome biogenesis. Annu Rev Cell Dev Biol. 2001;17:701-52 pubmed
- Draptchinskaia N, Gustavsson P, Andersson B, Pettersson M, Willig T, Dianzani I, et al. The gene encoding ribosomal protein S19 is mutated in Diamond-Blackfan anaemia. Nat Genet. 1999;21:169-75 pubmed
- Gazda H, Grabowska A, Merida Long L, Latawiec E, Schneider H, Lipton J, et al. Ribosomal protein S24 gene is mutated in Diamond-Blackfan anemia. Am J Hum Genet. 2006;79:1110-8 pubmed
- Cmejla R, Cmejlova J, Handrkova H, Petrak J, Pospisilova D. Ribosomal protein S17 gene (RPS17) is mutated in Diamond-Blackfan anemia. Hum Mutat. 2007;28:1178-82 pubmed
- Enenkel C, Lehmann A, Kloetzel P. Subcellular distribution of proteasomes implicates a major location of protein degradation in the nuclear envelope-ER network in yeast. EMBO J. 1998;17:6144-54 pubmed
- Materials and Methods [ISSN : 2329-5139] is a unique online journal with regularly updated review articles on laboratory materials and methods. If you are interested in contributing a manuscript or suggesting a topic, please leave us feedback.
- gene
- human ACOT8
- human AIF
- human AIM-1
- human AP2S1
- human ATG12
- human ATG5
- human B4GALT6
- human BinCARD
- human Bip
- human CBX1
- human CBX3
- human CBX5
- human CD3EAP
- human CD68
- human CD81
- human CD9
- human CENP C
- human CENPA
- human CLTB
- human COX-1
- human CPS1
- human Clathrin Heavy Chain
- human DCT
- human EEA1
- human FTCD
- human FUS
- human GLB1
- human GM130
- human GORASP1
- human H2AW
- human H2BC3
- human H3C7
- human H4C1
- human HK1
- human Hsc70
- human KDM1A
- human LAMP-1
- human LC3
- human Lamp-2
- human MLANA
- human NINL
- human NOP2
- human NUP98
- human PEX3
- human PMEL
- human PSMA1
- human PSMB5
- human PSMC1
- human PSMD1
- human PSMD7
- human RAB7A
- human RAB9A
- human RCAS1
- human RPL7A
- human RPS6
- human Rab11
- human Rab5
- human SEC13
- human SEC61B
- human TERF2IP
- human TGN46
- human TRF1
- human TUBA1A
- human TUBB3
- human TYRP1
- human VDAC1
- human calnexin
- human calreticulin
- human catalase
- human emerin
- human fibrillarin
- human gamma-adaptin
- human gamma-tubulin
- human giantin
- human lamin A/C
- human lamin B
- human mannosidase II
- human pericentrin
- human prohibitin
- human protein disulfide isomerase
- human ribosomal protein L26
- human smooth muscle actin
- human syntaxin 6
- human transferrin receptor