product summary
Loading...
company name :
Alomone Labs
product type :
chemical
product name :
α-Dendrotoxin
catalog :
D-350
more info or order :
citations: 33
| Reference |
|---|
Xu H, Guo W, Nerbonne J. Four kinetically distinct depolarization-activated K+ currents in adult mouse ventricular myocytes. J Gen Physiol. 1999;113:661-78 pubmed
|
Zhou B, Ma W, Huang X. Specific antibodies to the external vestibule of voltage-gated potassium channels block current. J Gen Physiol. 1998;111:555-63 pubmed
|
image
image 1 :
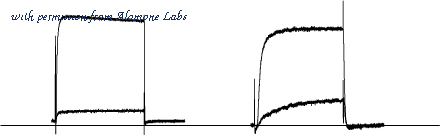
Alomone Labs ?-Dendrotoxin inhibits KV1.1 and KV1.2 channel currents heterologously expressed inXenopusoocytes. - KV1.1 (left in 2 mM K+) and KV1.2 (right in 5 mM K+) channel currents elicited by 200 ms depolarization from a holding potential of -100 mV to +20 mV before and during application of 100 nM?-Dendrotoxin(#D-350). 73% (n = 4) of the KV1.1 and 80% (n = 4) of the KV1.2 channels were inhibited by ?-Dendrotoxin respectively.
image 2 :

Western blot analysis of rat (lanes 1 and 3) and mouse (lanes 2 and 4)brain membranes: - 12.Anti-LINGO-1 (extracellular) Antibody (#ANT-032) (1:200).34. Anti-LINGO-1 (extracellular) Antibody preincubated with the negative control antigen.
product information
cat :
D-350
SKU :
D-350_0.14 mg
Product Name :
α-Dendrotoxin
Group Type :
Non Antibodies
Product Type :
Proteins
Accession :
P00980
Accession Number :
https://www.uniprot.org/uniprotkb/P00980/entry
Applications :
Electrophysiology
Formulation :
Lyophilized from double distilled water (ddH2O). May contain TFA as a residual counter ion.
Storage After Reconstitution :
The reconstituted solution can be stored at 4°C for up to 1 week. For longer periods (up to 6 months), small aliquots should be stored at -20°C. We do not recommend storing the product in working solutions for longer than a few days. Avoid multiple freeze-thaw cycles.
Reconstitution and Solubility :
Centrifuge the vial (10,000 × g for 5 minutes) before adding solvent to spin down all the powder to the bottom of the vial. The lyophilized product may be difficult to visualize. Add solvent directly to the centrifuged vial. Gently tap, tilt, and roll the vial to aid dissolution. Avoid vigorous vortexing; light vortexing for up to 3 seconds is acceptable if needed. The product is soluble in pure water at high micromolar concentrations (100 µM - 1 mM). For long-term storage in solution, we recommend preparing a stock solution by dissolving the product in double-distilled water (ddH2O) at a concentration between 100-1000x of the final working concentration. Divide the stock solution into small aliquots and store at -20°C. Before use, thaw the relevant vial(s) and dilute to the desired working concentration in your working buffer. Centrifuge all product preparations before use. It is recommended to prepare fresh solutions in working buffers just before use. Avoid multiple freeze-thaw cycles to maintain biological activity.
Solubility :
Centrifuge the vial before adding solvent (10,000 x g for 5 minutes) to spin down all the powder to the bottom of the vial. The lyophilized product may be difficult to visualize. Add solvent directly to the centrifuged vial. Tap the vial to aid in dissolving the lyophilized product. Tilt and gently roll the liquid over the walls of the vial. Avoid vigorous vortexing. Light vortexing for up to 3 seconds is acceptable if needed. The product is soluble in pure water at high micromolar concentrations (100 µM - 1 mM). For long-term storage in solution, we recommend preparing a stock solution by dissolving the product in double-distilled water (ddH2O) at a concentration between 100-1000x of the final working concentration. Divide the stock solution into small aliquots and store at -20°C. Before use, thaw the relevant vial(s) and dilute to the desired working concentration in your working buffer. Centrifuge all product preparations before use. It is recommended to prepare fresh solutions in working buffers just before use. Avoid multiple freeze-thaw cycles to maintain biological activity.
Storage Before Reconstitution :
The product is shipped as a lyophilized powder at room temperature. Upon receipt, store the product at -20°C. Protect from moisture.
Origin :
Dendroaspis angusticeps (Eastern green mamba).
Source :
Natural protein
Gene ID :
KCNA1 ,KCNA2 ,KCNA6
Product Page - Scientific background :
α-Dendrotoxin is isolated from Dendroaspis angusticeps snake venom by modification of the procedures of Harvey1 and Benishin2 and purified to homogeneity. α-Dendrotoxin blocks KV1.1 and KV1.2 channels (IC50= 0.4 to 4 and 1.1 to 12 nM in oocytes respectively, with higher values for mammalian cells).4 In addition, the toxin was shown to block KV1.6 (IC50= 9-25 nM).4α-Dendrotoxin was recently found to block ASIC currents in rat dorsal root ganglia with IC50 in the nM range5.
Supplier :
Alomone Labs
Target :
KV1.1, KV1.2, KV1.6, ASIC channels
Long Description :
A Blocker of KV1.1, KV1.2, KV1.6 and ASIC Channels
Short Description :
A Blocker of KV1.1, KV1.2, KV1.6 and ASIC Channels
MW :
7047 Da
Synonyms :
Venom basic protease inhibitor 1 homolog, Toxin C13S2C3, α-DTX
Modifications :
Disulfide bonds between: Cys7-Cys57, Cys16-Cys40 and Cys32-Cys53 Gln1 - Pyrrolidone carboxylic acid
Molecular formula :
C305H473N99O83S6
Effective Concentration :
10 - 500 nM
Activity :
α-Dendrotoxin inhibits 4-AP sensitive, inactivating voltage-gated K+ channels (KV1.1, KV1.2 and KV1.6).
Storage of solutions :
The reconstituted solution can be stored at 4°C for up to 1 week. For longer periods (up to 6 months), small aliquots should be stored at -20°C. We do not recommend storing the product in working solutions for longer than a few days. Avoid multiple freeze-thaw cycles.
Lead Time :
1-2 Business Days
Country of origin :
Israel/IL
Purity :
≥98% (HPLC)
CAS No :
74504-53-3
Form :
Lyophilized
Comment :
Contact Alomone Labs for technical support and product customization
Sequence :
QPRRKLCILHRNPGRCYDKIPAFYYNQKKKQCERFDWSG
CGGNSNRFKTIEECRRTCIG-OH
CGGNSNRFKTIEECRRTCIG-OH
Is Toxin :
Yes
UNSPSC :
12352202
Bioassay Tested :
yes
Steril endotoxin free :
no
Cited Application :
Electrophysiology
more info or order :
company information

Alomone Labs
Jerusalem BioPark (JBP), Hadassah Ein Kerem
P.O. Box 4287
Jerusalem 9104201
P.O. Box 4287
Jerusalem 9104201
info@alomone.com
http://www.alomone.com972 2 531 8002
headquarters: Israel
browse more products
questions and comments